
")
")
")
")
")
")
Kingdom: |
Animalia Chordata Mammalia Artiodactyla Ruminantia Cervidae Capreolinae Odocoileini Hippocamelus |
-
Hippocamelus bisulcus [Molina, 1782].
- Citation: Sagg. Stor. Nat. Chile, p. 320.
- Type locality: Chilean Andes, Colchagua Prov.
- Citation: Sagg. Stor. Nat. Chile, p. 320.
The taxonomic record (above) is taken from Wilson and Reeder (1993). No subspecies are recognized for the Patagonian huemul, although regional differences have been noted by various authors (e.g., Frid, 1994). This species is closely related to the taruca or North Andean huemul (H. antisensis), but the two are consistently recognized as separate species (Geist, 1998). Synonyms for Hippocamelus bisulcus include andicus, cerasina, chilensis, dubius, equinus, huemel, and leucotis (Wilson and Reeder, 1993).
Physical Characteristics
| Reported measurements for Patagonian huemul (Hippocamelus bisulcus) | ||||
| Source | Adult Weight | Head & Body Length | Shoulder Height | Tail Length |
| Diaz, 2000 | - | 163 cm 151 cm |
90 cm 81 cm |
13 cm |
| MacNamara, 1990 | 45-65 kg | 150-170 cm | 91 cm | 11.5-13 cm |
| Nowak, 1991 for Hippocamelus |
45-65 kg | 140-165 cm | 77.5-90 cm | 11.5-3 cm [sic] |
| Redford and Eisenberg, 1992 | 70.0 kg | 149.7 cm | 80.0-90.0 cm | 12-13 cm |
| Smith-Flueck, 2000 | 100 kg |
140-175 cm 140-157 cm |
79-100 cm 78-84 cm |
- |
| Whitehead, 1993 | 70-100 kg | - | 80-90 cm | - |
The coat is comprised of two layers: a long, greasy layer of bristle-like guard hairs which covers a dense, woolly undercoat (MacNamara, 1990; Nowak, 1991). The brittle guard hairs are hollow, which combined with the undercoat provides insulation to -50oC (MacNamara, 1990; Diaz, 2000). The pelage in winter is comprised of guard hairs 5-7 cm long; in spring they are shed in large clumps and replaced with a summer coat with hairs 3-4 cm in length (Diaz, 2000; Smith-Flueck, 2000). In all seasons, the hairs are slightly crinkled, growing the longest on the underside, tail, and legs (Nowak, 1991; Smith-Flueck, 2000).
The build of the Patagonian huemul is distinctly stocky, a result of an arched back and relatively short legs (Nowak, 1991; Smith-Flueck, 2000). The resulting crouching position is typical of many mountain-dwelling ungulates, as are the hard, pointed, sharply-edged hooves (Geist, 1998; Smith-Flueck, 2000). Tarsal glands are present on the rear legs, but metatarsal glands are absent (Geist, 1998).
The ears are long and narrow (described by MacNamara (1990) as "donkey ears"); they measure around 17 cm in length, and are covered on their inner surface with white hair (Diaz, 2000; Smith-Flueck, 2000). The only facial marking is a dark forked blaze on ridge of the nose, which runs from the dark rhinarium and splits in between the eyes to form 'eyebrows' (Smith-Flueck, 2000). While this blaze is usually attributed to males alone, both sexes possessed such markings in a coastal Chilean population studied by Frid (1994). The eyes are surrounded by a vague area of lighter colored hair, which accentuates the large preorbital glands (Geist, 1998; Diaz, 2000). Varying authors report either a presence (MacNamara, 1990; Nowak, 1991) or absence (Smith-Flueck, 2000) of upper canine teeth.
Only males grow antlers; these are usually simple in form and typically branch just once, although antlers with up to five tines have been recorded (Nowak, 1991; Redford and Eisenberg, 1992; Smith-Flueck, 2000). In a typical forked antler, the rear tine is longer, typically measuring 30-35 cm in length from the burr (where the antler joins the skull) to the tip of the tine (Nowak, 1991; Redford and Eisenberg, 1992, Diaz, 2000). The first set of antlers in young males appears around eighteen months of age, although they may appear as early as five months of age (Redford and Eisenberg, 1992; Smith-Flueck, 2000). Males cast off their antlers near the end of the austral winter, from mid July to early September (Smith-Flueck, 2000), although Geist (1998) states that they are dropped in October-November. New antlers begin growing soon after old ones are shed, and the full size is reached (and velvet shed) after around five months of growth (Smith-Flueck, 2000).
Reproduction and Development
The gestation period has never been measured directly, but estimates range from 6-8 months - reported numbers include 180-210 days (Redford and Eisenberg, 1992), 200-220 days (Smith-Flueck, 2000) and 240 days (Geist, 1998). Females almost always give birth to a single young, usually in the months of November and December, a time when the local vegetation is highly nutritious (Frid, 1994; Smith-Flueck, 2000). Fawns are born with a thick, woolly coat which is not spotted (unlike many deer species), but rather a solid dark coffee-brown color (MacNamara, 1990; Redford and Eisenberg, 1992; Smith-Flueck, 2000). A single newborn huemul examined by Smith-Flueck (2000) weighed 2035 g, with a total body length of 61.5 cm. Milk teeth show at birth (Smith-Flueck, 2000).
Prior to giving birth, females seek seclusion; born in an isolated area (often along high bluffs), the newborn fawn will spend most of its time bedded down and separated from its mother, with the resting site being changed frequently to avoid predators. Eventually, the fawn will begin to follow its mother, although at a stomp of the dam's feet (indicating danger) the fawn immediately drops to the ground and freezes. Weaning occurs at 4-5 months of age (Smith-Flueck, 2000).
Despite the fact that females may reach sexual maturity as early as 6 months of age, the fecundity of H. bisulcus is low, with fewer than 50% of females breeding on an annual basis. Smith-Flueck (2000) suggests that this may be due to a long interbirth interval, resulting in females only breeding every other year. The maximum observed life span for this species is around 14 years of age (Smith-Flueck, 2000).
Ecology
Huemuls are frequently observed on north or north-western facing slopes, often with a pitch between 30 and 40 degrees (MacNamara, 1990; Redford and Eisenberg, 1992; Smith-Flueck, 2000). In Chubut, Argentina, however, observations of this species have been made on slopes generally less than 25 degrees (Smith-Flueck, 2000). Patagonian huemul are usually found at or below the tree line, principally inhabiting brushy areas with dense shrub cover (65% of observations in Argentina) and temperate forests (20% of observations in Argentina) (MacNamara, 1990; Nowak, 1991; Redford and Eisenberg, 1992; Smith-Flueck, 2000). These subalpine forests are dominated by false-beeches Nothofagus sp. (including N. antarctica, N. betuloides, and N. pumilio; Fagaceae), with shrub cover including Discaria sp. (Rhamnaceae), Gaultheria sp.(Ericaceae), Embothrium coccineum (Proteaceae), Escallonia rubra (Escalloniaeae), and Mulinum spinosum (Umbelliferae) (MacNamara, 1990; Galende et al., 2005).
Seasonal movements are well-documented in this species: huemuls inhabit high altitudes during summer, with frequent usage of pasture and brush above the treeline (80%) or in forests (12.5%) (Nowak, 1991; Smith-Flueck, 2000). Females especially prefer high cliffs and ridges (likely for the protection they offer from terrestrial predators), while males are found almost exclusively on flatter meadows (Frid, 1994; Smith-Flueck, 2000). From these high elevations, huemuls move downslope in autumn in order to winter in valleys and denser forests (Nowak, 1991; Redford and Eisenberg, 1992). Altitudinal variations of two to three kilometers were documented for one animal (Smith-Flueck, 2000).
Local population density estimates range from 0.02 to 5.66 animals per square kilometer (Smith-Flueck, 2000), although Redford and Eisenberg (1992) report only the lower end of this range (0.02 to 1.2 per square kilometer). Annual home range size in Tamango Nature Reserve, Chile was estimated to be 300 hectares (Smith-Flueck, 2000), a figure which compares well to the 340 hectares reported in Redford and Eisenberg (1992). During the breeding season (between March and May), huemuls confine their movements to ranges of 36 to 73 hectares, although this is likely a gross underestimate of the area used throughout the year (Smith-Flueck, 2000). On a daily basis, individuals may travel 3.0-3.9 kilometers as they search for food and shelter (Smith-Flueck, 2000).
Predators of the huemul include the puma (Puma concolor) and domestic dogs (Canis lupus familiaris), while the culpeo fox (Pseudoalopex [Dusicyon] culpaeus), wild cats (Oncifelis guigna and O. geoffroyi), and raptors (especially condors) may be a threat to young (Geist, 1998; Smith-Flueck, 2000). When threatened by predators, H. bisulcus attempts avoidance by first hiding, but if discovered at close range will flee rapidly (Geist, 1998). This species has low endurance when running, and will escape persistent pursuers by seeking refuge on cliffs or by swimming into lakes: this species swims with ease (Geist, 1998; Diaz, 2000; Smith-Flueck, 2000).
H. bisulcus is a browser, feeding almost exclusively on tender vegetation and only rarely on grasses or lichens (Geist, 1998; Smith-Flueck, 2000). Several studies on food consumption have been conducted, which show a strong trend with some regional variation. In addition to the species listed below, captive animals have shown a preference for willow (Salix sp.) (Smith-Flueck, 2000).
Galende et al. (2005), observing huemul in Nahuel Huapi National Park, identified 32 plant species consumed by H. bisulcus. Shrubs and trees comprised the greatest part of the diet (58.6% and 19.5% of fragments identified in feces, respectively), while forbs and grasses were occasionally consumed, each contributing to just over 9% of the diet. The shrub Ribes magellanicum (Grossulariaceae) and the tree Nothofagus pumilio (Fagaceae) were the principal components of the diet during the summer (28.5% and 33.4%, respectively), while in winter (when these plants drop their leaves) the shrubs Maytenus sp. (Celastraceae) and Gaultheria sp. (Ericaceae) were used heavily (50.8% and 28.7%, respectively).
Smith-Flueck (2000) reports 48 plant species which have been identified in the diet, comprised of 27-46% forbs and 31-72% woody plants when averaged across the year. Maytenus were a very important food item, making up as much as 60% of the annual diet in Argentina. Other frequently used species include Alstroemeria (9%), Chlorea viridiflora (Orchidaceae; 8%), Geranium (5%), and Schinus (5%).
The population living along the Chilean coast observed by Frid (1994) had a more restricted diet, with only 11 species of plant being eaten. The perennial forb Gunnera magellanica (Gunneraceae) was heavily preferred, with consumption of this plant accounting for as much as 82.5% of observed feeding time in grasslands and bluffs. Feeding in moorland habitat was concentrated on two species: the tree Nothofagus antarctica (Fagaceae; 84.7% of observation time) and evergreen shrub Embothrium coccineum (Proteacea; 15.3% of observation time).
Behavior
Patagonian huemuls are either solitary or found in small groups; these are generally composed of 2-5 individuals, although aggregations of up to ten individuals have been observed at low altitudes (Redford and Eisenberg, 1992; Smith-Flueck, 2000). The structure of these groups is highly flexible, and a single individual may associate in several 'groups' during the course of a single day (Smith-Flueck, 2000). Adults of both sexes are often seen alone, while the most commonly observed grouping is a mother-offspring pair (29%). Pairs of an adult male and female, either alone or accompanied by juveniles, are also seen regularly, while groups of adult males are infrequent and all-female groups never observed. Adult males tolerate the presence of subordinate males, and it has thus been inferred that territoriality is absent in this species (Smith-Flueck, 2000).
The courtship behavior of the Patagonian huemul is similar to other New World deer, and has been described in detail by Smith-Flueck (2000). As the breeding season approaches, males thrash their antlers against vegetation, helping to shed the velvet from the fully formed antlers while advertising their presence (they may fray the bark off of tree trunks from just above the ground up to 113 cm high). This behavior continues through the courtship period, and is coupled with scent marking using the facial glands (Nowak, 1991). A male huemul will follow a potentially receptive female sniffing her anogenital region and employing flehmen (lip-curl) after smelling her urine in order to determine whether or not the female is in estrus. After determining the reproductive status of a female, the male will approach receptive mates with the head stretched low, performing a high amount of "foreplay" and non-copulatory mounting before achieving intromission. Although a male (buck) will mate with many females (does) as they become receptive, the association with a single doe may last for as long as 8-9 weeks, including a long period of time after mating. Dominant males may chase off subordinates if they approach within 300 meters of an estrus female (Geist, 1998). Otherwise, the low population density at which huemuls live minimizes the amount of conflict between males, and may explain the reduced antlers of this species as well as the low frequency of dominance displays (Smith-Flueck, 2000).
Smith-Flueck (2000) describes this species' repertoire of vocalizations, many of which are audible to humans at a distance of 30 meters or more. During the breeding season, males either a grunt or produce a series of soft whines while pursuing females; a doe, in return, will produce a snort when she is the object of a male's attention. Agonistic encounters between males are accompanied by a rapid series of sharp notes, lasting 3-6 seconds and descending in volume throughout the series; Smith-Flueck (2000) suggests that older males may produce deeper noises, thereby advertising the male's status. The alarm call of the Patagonian huemul is a snort, which is usually accompanied by the stamping of the front legs. Fawns bleat in alarm; these cries usually bring the mother immediately to the fawn.
Genetics
Distribution
Countries: Argentina, Chile (Jiménez et al., 2008).
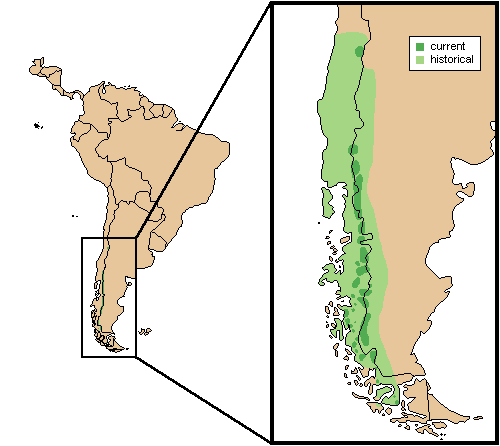
Conservation Status
Threats to huemuls are both ongoing and numerous. Humans affect the huemul directly through hunting (poaching) and by encroaching on this species' habitat (Jiménez et al., 2008). In the Nevados area of Chile (the site of the most northerly population of huemul), land settlement, construction of pipelines, and logging (along with the construction of logging roads) have all served to reduce native habitats and increase the presence of humans in the region (Povilitis, 1998). An influx of tourists has also occurred, although the impacts of tourism have yet to be determined (Flueck and Smith-Flueck, 2006). However, perhaps the largest threat comes from domestic stock and introduced species (Redford and Eisenberg, 1992; Jiménez et al., 2008). Harassment and depredation by domestic dogs, susceptibility to diseases and parasites of introduced species (especially domestic cattle and sheep), and competition from introduced species (e.g., red deer) are all major factors in the continued decline of this species (Povilitis, 1998; Flueck and Smith-Flueck, 2006; Jiménez et al., 2008)
This species is actively protected in Chile, and a conservation agreement between Argentina and Chile is in effect (Weber and Gonzalez, 2003). However, Flueck and Smith-Flueck (2006) expressed concern regarding the actual conservation status in Argentina; these authors advocate increased conservation measures be taken if this species is to continue to survive. In 2007, the Huemul Task Force was created under the IUCN Species Survival Commission to provide recommendations - based on sound scientific information - through which the recovery of huemul can be achieved.
Remarks
The common name huemul (pronounced hw~y~mul) is of Araucanian origin, and is said to derive from the verb "huemin", meaning "to follow"; this is apparently in reference to a custom of this species of walking in single file (MacNamara, 1990; Smith-Flueck, 2000). The name is also occasionally spelled as guemal. In the past, this species was popularly known as the "Chilean huemul". However, since many populations inhabit Argentina, the use of this name has been replaced by the more appropriate name "Patagonian huemul" (Smith-Flueck, 2000). An increase in the use of the local name "taruca" for H. antisensis has meant that H. bisulcus is now sometimes referred to simply as the "huemul".
The genus name Hippocamelus is a combination of the Greek words hippos (a horse) and kamelos (a camel) - while huemuls are not closely related to either, they were first described (from hearsay information) as a type of wild ass (Diaz, 2000). The species name bisulcus translates from Latin as "having two furrows", or "cloven"; which Diaz (2000) suggests refers to original classification of this deer as a cloven-hoofed horse.
-
- Local names
- Huemul [from Araucanian Indians] (MacNamara, 1990; Smith-Flueck, 2000)
-
- French
-
Huémul des Andes méridionales (Jiménez et
al., 2008)
- Huemul du Chili (MacNamara, 1990)
-
- German
- Südlicher Andenhirsch (MacNamara, 1990)
-
- Spanish
-
Huemul Patagónico (Redford and Eisenberg, 1992)
Ciervo Andino meridional (Jiménez et al., 2008)